Ottica Campagnacci
Ottica Campagnacci










Fabrizio Campagnacci

www.otticacampagnacci.com
/home/blog

OTTICA CAMPAGNACCI aderisce alla raccolta degli occhiali usati:





OTTICA
CAMPAGNACCI
is authorized dealer of:

- Binocoli e cannocchiali
- Microscopi
- Strumenti elettronici meteo
- Altimetri e bussole
- Barometri, termometri e Galileiani
- Lenti d'ingrandimento
- Telescopi
- Visori notturni "Yukon"
- Strumenti "Suunto"
- Catalogo "C.A.I."



Meccanismo della visione

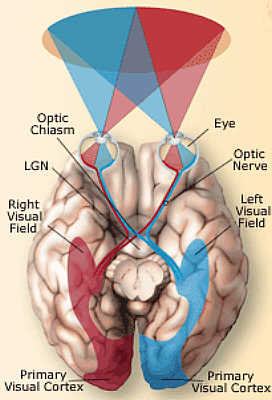
La percezione visiva avviene attraverso una serie di processi che implicano non solo fattori fisiologici, ma anche una complessa rielaborazione dei segnali luminosi da parte del cervello.
La prima tappa di questi processi ha sede negli occhi: essi sono responsabili della ricezione e traduzione dell’energia in un segnale nervoso. Questo segnale viene poi elaborato nella corteccia
cerebrale, dove si attivano i neuroni sensibili alle varie caratteristiche fisiche e cognitive degli stimoli. In realtà è solo a questo punto che si può parlare di percezione perché gli stimoli
cominciano ad acquisire un significato solo una volta elaborati dal nostro cervello.
Per definire in maniera completa il processo della visione occorre quindi esaminare le funzioni dei vari organi e come questi interagiscano fra loro.
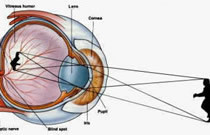
Fisiologia dell'occhio
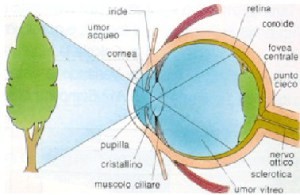
L’occhio è un organo complesso che racchiude sia il sistema ottico vero e proprio, che permette il formarsi delle immagini sulla retina, sia una propaggine del sistema nervoso centrale, che
opera una prima analisi di tali immagini.
Il suo funzionamento è assimilabile a quello di un comune sistema ottico (come quello di una macchina fotografica o di un canocchiale) costituito da un diottro (cornea, umor acqueo, e umor
vitreo) di indice di rifrazione 1.33 e da una lente biconvessa, il cristallino, di indice di rifrazione 1.44.
Il suo funzionamento è assimilabile a quello di un comune sistema ottico (come quello di una macchina fotografica o di un canocchiale) costituito da un diottro (cornea, umor acqueo, e umor vitreo) di indice di rifrazione 1.33 e da una lente biconvessa, il cristallino, di indice di rifrazione 1.44.
Il suo funzionamento è assimilabile a quello di un comune sistema ottico (come quello di una macchina fotografica o di un canocchiale) costituito da un diottro (cornea, umor acqueo, e umor vitreo) di indice di rifrazione 1.33 e da una lente biconvessa, il cristallino, di indice di rifrazione 1.44.
La cornea è una sottile pellicola trasparente che chiude anteriormente l’occhio ed ha forma sferica. Questa calotta è assimilabile ad una lente a
menisco con potenza di 42 diottrie (ricordio che una diottria è l’inverso di un metro), in grado di assorbire le nocive radiazioni ultraviolette: in questo modo protegge le parti interne dal
danneggiamento che queste radiazioni potrebbero produrre. Il difetto dell’astigmatismo si presenta allorchè, anzichè sferica, la cornea presenta una forma elissoidale.
L’umor acqueo, come dice la parola, è un liquido salino interposto tra la cornea e il cristallino ed ha lo scopo, mediante una lieve pressione sulla parete
interna della cornea, di mantenerne la forma. Anche l’umor vitreo è una sostanza gelatinosa che riempie totalmente l’occhio in modo da fargli mantenere una
forma sferica.
Il cristallino funziona come una lente biconvessa con curvature differenti ed è costituito da diversi strati sovrapposti a guisa di cipolla, la cui
potenza è di circa 20 diottrie; purtroppo con l’età o tramite agenti ionizzanti (come un intenso irraggiamento ultravioletto) tende a opacizzarsi e quando l’opacizzazione è tale da
compromettere una visione distinta degli oggetti (cataratta) se ne rende necessaria la rimozione. Il cristallino è connesso a fibre muscolari che gli permettono di variare la propria curvatura in
modo da far cadere costantemente sulla retina il piano focale dell’immagine (accomodamento del cristallino); col passare degli anni purtroppo perde questa sua proprietà, facendo insorgere il
fenomeno della presbiopia.
Combinando la potenza della cornea e quella del cristallino si ottiene una potenza totale dell’occhio pari a 60 diottrie; cornea, camera anteriore, cristallino e camera posteriore nel loro
complesso formano una lente convergente (provvista di una distanza focale variabile fra 2,4 e 1,7 cm) che proietta le immagini sulla retina, rimpicciolite e capovolte.
Una membrana muscolare, l’iride, al cui centro e’ ricavata un’apertura, la pupilla, e’ quella struttura che
tipicamente porta il colore dei nostri occhi e serve a diaframmare, cioe’ a regolare la quantita’ di luce che entra nell’occhio: in presenza di poca luce il foro tenderà a dilatarsi (fino a circa
8 mm in una persona giovane), mentre in condizioni di alta luminosità tenderà a contrarsi fino a 2 mm. La condizionione migliore per la visione corrsiponde a una dilatazione intermedia: una
pupilla troppo chiusa può dar luogo a problemi di diffrazione mentre una pupilla troppo dilatata evidenzierà le aberrazioni del sistema ottico.
La retina
La retina è giustamente ritenuta la parte più importante dell’organo visivo. Essa tappezza interiormente tutto l’occhio ed è una struttura assai complessa.
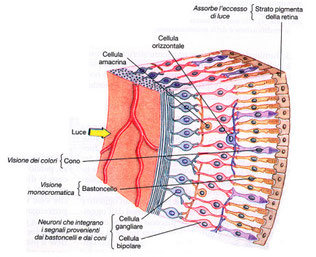
Come si vede in figura, partendo dalla parte più lontana dalla cornea troviamo lo strato
deifotorecettori (coni e bastoncelli), lo strato delle cellule
orizzontali, bipolari, amacrine e gangliari, e il nervo ottico. Il compito dei fotoricettori è quello di trasformare in
impulsi elettrici le informazioni ricevute dalle reazioni fotochimiche che vengono attivate dalla radiazione luminosa e di inviare questi segnali ai neuroni retinici – le cellule orizzontali,
bipolari, amacrine e ganglionari – che sono variamente connessi fra di loro ed effettuano una prima elaborazione del segnale visivo.
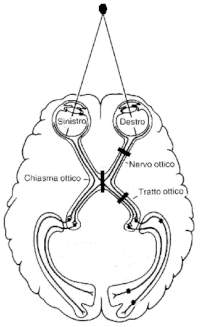
Gli assoni delle cellule gangliari si riuniscono in modo da formare il nervo ottico, un cavo che conduce l’informazione visiva fuori dalla retina fino ai centri superiori, dapprima al corpo
genicolato laterale e da qui alle aree corticali.
I coni, più grossi e più corti dei bastoncelli, se ne differiscono anche come struttura, mentre il loro numero ne è notevolmente inferiore: nell’occhio umano sono presenti circa 115 – 120 milioni
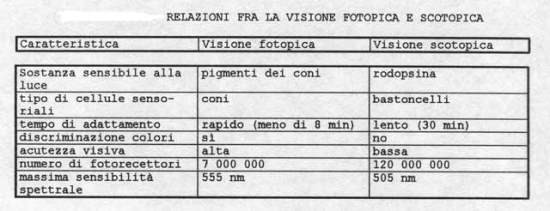
di bastoncelli contro i 6 – 7 milioni di coni. I coni sono demandati alla visione diurna (fotopica), mentre i bastoncelli, per la loro maggiore sensibilità, sono preposti alla visione notturna
(scotopica).
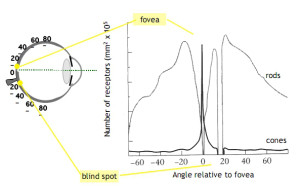
La distribuzione dei coni e dei bastoncelli varia sulla retina: la loro concentrazione (e di conseguenza l’acuità visiva) diventa molto grande in una zona che si presenta come una depressione
rispetto alla circostante retina, di diametro circa 2mm, la macula lutea, così chiamata perché gli strati maculari interni sono permeati da un pigmento giallognolo.
Al centro della macula lutea si trova una zona, la fovea centralis, di diametro circa 0.25 mm, contenente praticamente solo coni, al massimo della loro concentrazione (circa
il 90 % della loro totalità sulla retina). Questa si presenta come la massima depressione rispetto alla restante retina ed è la zona ottimale per la formazione dell’immagine: l’occhio infatti
ruota nella sua orbita allo scopo di focalizzare l’immagine degli oggetti in questa zona. Man mano che ci si allontana dalla fovea il numero dei coni diminuisce fino ad annullarsi nella zona
periferica della retina, dove si trovano solo bastoncelli.
Dalle immagini si vede come la luce arriva allo strato dei fotorecettori solo dopo aver attraversato l’intero spessore della retina, eccezion fatta per la fovea: in questo punto la altre cellule
retiniche si ritraggono, lasciando esposti soltanto i fotorecettori (principalmente coni).
Oltre alla fovea, esiste anche un altra zona,di circa 1,5 mm di diametro, che si differenzia dalle altre: in questo punto convergono i nervi e i vasi sanguigni della retina e pertanto
quest’area, che viene chiamata “punto cieco”, non è sensibile alla luce. Tuttavia generalmente non ci accorgiamo di questo buco, perchè il cervello riesce a ricostruire
l’immagine mancante attraverso un processo chiamato “filling in”.
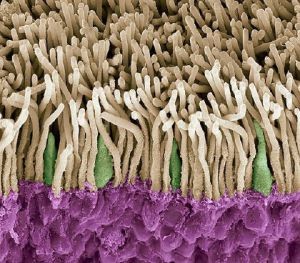
I coni e i bastoncelli
Come accennato sopra, nella parte più esterna della retina sono situate le cellule sensibili alla luce (fotorecettori), i coni (in verde nella foto sottostante) e i bastoncelli (in bianco nella foto sottostante).
Nei fotorecettori avviene la fototrasduzione, ovvero il processo attraverso il quale il segnale luminoso viene convertito in segnale nervoso. La regione dei fotorecettori che contiene
i fotopigmenti, ovvero le molecole che hanno la funzione di assorbire la luce, si trovano nel segmento esterno dei fotorecettori, che è cilindrico nei bastoncelli ed appuntito nei coni (da cui
appunto il nome dei due tipi di fotorecettori).
I fotopigmenti (o opsine), a contatto con un fotone, cambiano la propria struttura molecolare scatenando una iperpolarizzazione che rende fortemente negativo il potenziale di membrana:
l’iperpolarizzazione determina una diminuzione nella produzione di glutammato da parte del fotoricettore, alterazione che determina la produzione di stimoli nervosi visivi.
I bastoncelli risultano più sensibili alla luce rispetto ai coni, grazie al loro pigmento visivo, la rodopsina, presente in maggiore quantità.
Possiedono inoltre una risposta lenta al fotone di luce: in seguito all’assorbimento di un fotone, la risposta in termini elettrici del bastoncello dura molto più a lungo di quanto risulti essere
l’esposizione vera e propria del bastoncello alla luce; nel cono invece questo non accade e la risposta è molto più veloce. Per questo motivo quando ci rechiamo in una stanza buia, la
sensibilità dei fotorecettori aumenta molto lentamente (fenomeno dell’adattamento), raggiungendo il massimo dopo circa 30 min (visione scotopica mediata soprattutto dai bastoncelli).
Di contro i bastoncelli possiedono un’alta amplificazione del segnale e sono in grado di rispondere, al massimo della loro sensibilità (visione scotopica), addirittura al singolo fotone di
luce, in virtù del tempo di integrazione della risposta così lungo.
Questo tipo di fotorecettori presenta inoltre una risposta che si chiama saturante: oltre una certa soglia luminosa, tutte le molecole di rodopsina
subiscono una variazione della propria struttura ed ulteriori stimoli luminosi non scatenano più alcuna risposta (accecamento).
Da quanto detto emerge che i bastoncelli presentano una elevata sensibilità, tanto da riuscire a rispondondere al singolo fotone nelle condizioni ottimali; il rovescio della medaglia si
riscontra nella saturazione della risposta a stimoli troppo intensi. Quindi funzionano al loro meglio quando il livello di luminosità generale è molto basso, grazie al fatto che possono
amplificare enormemente la poca luce presente.
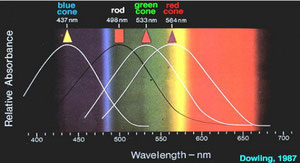
Infine, possedendo un solo fotopigmento, i bastoncelli possono fornire una visione unicamente monocromatica: la rodopsina assorbe a 498 nm, quindi la luce più efficiente a stimolare i
bastoncelli è sostanzialmente compresa fra il blu e il verde (vedi figura sottostante).
I coni non sono sensibili come i bastoncelli: al massimo della loro sensibilità hanno bisogno di almeno un centinaio di fotoni per iniziare a
rispondere alla luce; in compenso hanno una risposta più veloce di quella dei bastoncelli e saturano per quantità di luce molto più alte di quelle per cui saturano i bastoncelli.
Quindi, se la luce ambientale è scarsa ci serviremo della grande sensibilità dei bastoncelli per vedere mentre se la luce ambientale è elevata i bastoncelli saranno saturati e ci serviremo
dei coni.
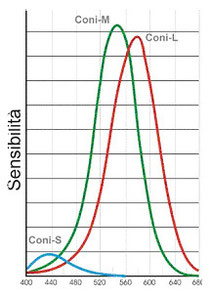
I coni, al contrario dei bastoncelli, possiedono 3 tipi di pigmenti visivi che mediano le informazioni sui colori che noi percepiamo:(ogni cono della retina contiene solo uno dei tre
fotopigmenti): i coni cosiddetti “blu“ (short) assorbono prevalentemente la luce a 437 nm, che è nel blu-violetto appunto, come
evidenziato nella figura sopra; i coni cosiddetti “verdi”(middle) hanno un picco di assorbimento della luce a 533 nm, corrispondente a un
azzurro-verde; i coni cosiddetti “rossi“ (long) hanno in realtà un picco d’assorbimento a 564 nm che, di fatto, è un colore
giallo-verde.
Ovviamente, i pigmenti non sono colorati, ma il nome è stato loro dato in base alla sensazione associata alla loro massima sensibilità.
La sensazione di colore è legata ai rapporti delle attività neurali dovute ai tre fotopigmenti: ad esempio un raggio luminoso monocromatico della lunghezza d’onda di 450 nm produce una
risposta forte dal fotopigmento “blu”, una risposta più debole dal “verde” ed una più debole ancora
dal “rosso”. Inoltre i diversi tipi di coni non sono distribuiti uniformememente: il 60% circa dei coni contiene il pigmento “rosso”, mentre
il 30% contiene il “verde” e solo il 10% contiene il pigmento “blu”.
Dalla figura si nota come le sensibilità assolute dei tre diversi tipi di coni sono molto differenti fra di loro: questo comporta una maggiore sensibilità verso la zona giallo-verde
dello spettro luminoso in ambito fotopico.
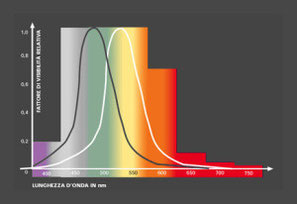
Visione fotopica, mesopica e scotopica
Per i motivi espressi nel capitolo precedenti, la CIE (Commision Internationale de l’Eclairage) ha definito rispettivamente nel 1924 e nel 1955 due curve normalizzate che rappresentano il
fattore di visibilità relativo in funzione della lunghezza d’onda per visione fotopica e scoptopica.
Per la visione fotopica tali valori sono rappresentati dalla funzione V(λ) (indicata nello schema seguente in bianco) che misura l’efficienza
visuale a varie lunghezze d’onda nel caso di luminosità diurna.
Essa è normalizzata al massimo valore V(λ)=1 relativo alla lunghezza d’onda di 555,02 nm. Questa funzione, basata su un campione di
circa 200 persone, è stata internazionalmente accettata, anche se recentemente vi è molta discussione sulla sua
effettiva correttezza.
Una differente funzione V’(λ) (indicata nello schema in nero) misura l’efficienza dell’occhio nel caso di un livello di luce inferiore, tipico della visione
scotopica. In queste condizioni il valore V’(λ)=1 si ha per la lunghezza d’onda di 507 nm. Lo spostamento del massimo di sensibilità, dovuto all’utilizzo, da parte
dell’occhio, prima di coni e poi di bastoncelli, è denominato effetto Purkinije. I bastoncelli, che
funzionano in condizioni di bassa visibilità, vedono meglio il blu di quello che fanno i coni, i quali
possono vedere luce profondamente rossa, luce che per i bastoncelli appare nera.
Se si hanno due pezzi di carta colorata rossa e blu, in condizioni di buona luminosità, risulta più luminoso il pezzo rosso, passando
all’oscurità l’effetto si inverte.
La visione mesopica è la visione dovuta all’attività contemporanea dei bastoncelli e dei coni della retina. Si tratta del tipo di visione che si ha
quando il livello di illuminazione è intermedio (definito generalmente dall’intervallo di luminanza compreso fra 0,001 e 3 cd/mq).
Acuità visiva
L’ acuità visiva (o acutezza visiva) è la capacità dell’occhio di risolvere e percepire dettagli fini di un oggetto e rappresenta l’inverso delle
dimensioni angolari minime che un oggetto deve avere per poter essere percepito correttamente.
Il minimo angolo di risoluzione corrisponde al minimo angolo visivo che separa due punti per cui i due punti appaiono ancora separati; se mi allontano da loro l’angolo visivo scenderà sotto
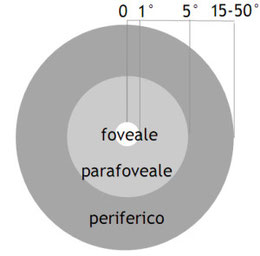
l’angolo limite ed io non vedrò due punti separati ma un punto solo. Nell’uomo l’acuità visiva foveale è di 1’, ovvero un sessantesimo di angolo visivo, mentre è molto minore per le altre
regioni.
Per spiegare queste differenze occorre innanzitutto evidenziare come i bastoncelli (presenti soprattutto nelle aree perfieriche della retina,
come evidenziato in precedenza) hanno una via di comunicazione molto convergente: questo significa che centinaia di bastoncelli, o migliaia di bastoncelli nelle regioni più periferiche della
retina, convergono su un’unica cellula ganglionare. Questo vuol dire che un singolo assone convoglierà le informazioni che raccolgono un numero molto elevato i bastoncelli, distribuiti su di
un’area estesa della retina, con conseguente perdita di moltissima informazione spaziale.
Di conseguenza il campionamento spaziale diventa grossolano, informazioni raccolte in punti diversi della retina sono sommate insieme.
La via di comunicazione dei coni è invece poco convergente: a livello della regione centrale della fovea si raggiunge addirittura un rapporto 1:1 tra
cono e cellula ganglionare; in questo punto otteniamo la migliore risoluzione spaziale grazie inoltre al minore spessore della retina, che consente ai fotoricettori di essere direttamente a
contatto con i segnali luminosi.
Nella fovea sono presenti solo coni che sono densamente impacchettati in un mosaico ordinato che raggiunge la densità di 160.000 coni/mmq e la distanza fra una fila di coni e quella
adiacente è di 2.5 mm: in questa zona il mosaico dei coni foveali funziona al massimo delle sue possibilità; è del resto abbastanza intuitivo che un campionamento fitto (molti recettori/mmq)
consente di rilevare meglio i dettagli dell’immagine di un campionamento rado.
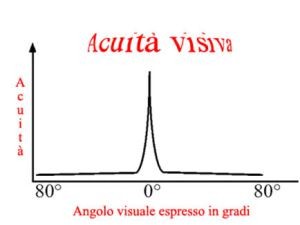
Nella figura a destra si osserva che, procedendo dalla fovea verso la periferia della retina, e quindi andando verso zone in cui la densità dei coni diminuisce drammaticamente,
l’acuità visiva decresce rapidamente: a 10 gradi di eccentricità è già diminuita di un fattore 10, in ottimo accordo con la riduzione nella densità dei coni. L’aumento di densità dei bastoncelli
verso le zone periferiche non riesce a sopperire alla diminuzione dei coni in termini di acuità visiva perchè, come detto in precedenza, molti bastoncelli convergono su una stessa cellula
gangliare.I bastoncelli quindi non sono quindi un sistema ad alta acuità visiva: del resto, siamo tutti consapevoli che tentare di leggere con la periferia della retina o in condizioni di
luce scarsa, ovvero usando il sistema dei bastoncelli, non dà buoni risultati!
I coni foveali, invece, a causa del loro fitto impacchettamento e dell’assenza di convergenza ci consentono un’alta acuità visiva, purché la luminosità sia sufficiente per il loro
funzionamento.
Alla luce di quanto detto, la visione foveale (o visione distinta) è caratterizzata dalla sensibilità al colore, dalla massima acuità visiva, dalla discriminazione e
riconoscimento dell’oggetto, unita però alla mancanza di sensazione del movimento.
La visione periferica è invece caratterizzata da insensibilità al colore, da bassa acuità visiva e dalla capacità di avvistamento (ma non discriminazione e
riconoscimento) di oggetti presenti nello spazio circostante, unito a funzioni di sorveglianza, scoperta, avviso ed in particolare alla sensazione del movimento.